|
Все последние события из жизни вулканологов, сейсмологов |
|
|
2018-01-01 09:00 Есть такая профессия – метан окислять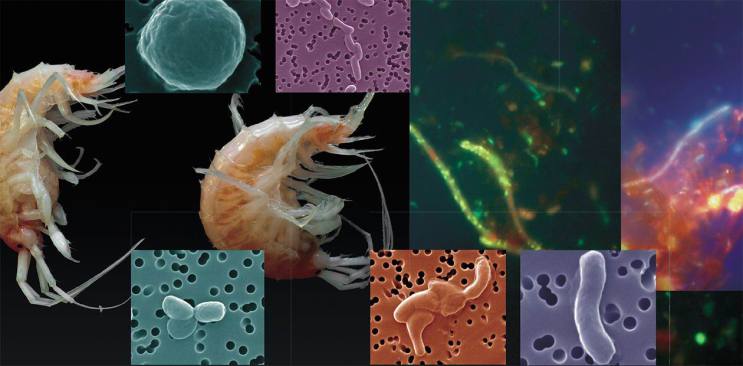
Есть такая профессия – метан окислять
На планете человек сосуществует с огромным, невидимым глазу миром микроорганизмов – царством бактерий. В этом царстве все как у людей: бактерии дружат «по интересам», а некоторые предпочитают жить по принципу «ты мне – я тебе». Специальностей у бактерий много, и все они профессионалы в своей области. Например, метанотрофы «едят» метан и могут жить в самых разных, в том числе экстремальных, условиях среды. Метанокисляющие организмы были обнаружены и в микробных сообществах «грязевых вулканов» в тектонически активных зонах Байкала рядом с глубинными выходами метана Метан – это предельно восстановленное органическое соединение, недоступное для живых организмов. Исключение составляют метанокисляющие бактерии (метанотрофы) – уникальные биологические системы, которые используют метан в качестве единственного источника энергии и углерода. Окисляя этот простейший углеводород, они производят углекислый газ – «пищу» для фотосинтезирующих растений. Метанотрофы обнаруживаются в самых разных экосистемах, поскольку основными факторами, определяющими их жизнедеятельность, служит наличие метана и кислорода. Источников же метана в биосфере много. Основными естественными «поставщиками» метана являются северные увлажненные экосистемы – болота и тундра с заболоченными почвами, а также осадочные отложения морей, озер, прудов и т.?п. Много метана выделяют рисовые поля, термитники и, как ни удивительно, крупный рогатый скот. Среди антропогенных источников метана – свалки и разрабатываемые месторождения каменного угля и природного газа. Важная роль метана в биосфере Земли определяется его способностью поглощать инфракрасное излучение, отраженное от поверхности планеты. И хотя его концентрация в атмосфере невелика, он занимает второе место после углекислого газа в списке так называемых «парниковых газов», влияющих на состояние климата. В этом смысле метанотрофы являются своего рода биологическим фильтром, препятствующим избыточной эмиссии метана в атмосферу. В вечной мерзлоте и желудке коровы Многоступенчатый процесс окисления метана у метанотрофов осуществляется благодаря особой ферментной системе. Первичную «атаку» молекулы субстрата осуществляет сложный ферментный комплекс – метанмонооксигеназа. Образующиеся затем промежуточные продукты реакции (метанол, формальдегид и формиат) последовательно окисляются специфическими ферментами. Ассимиляция формальдегида у разных метанокисляющих бактерий происходит в разных биохимических циклах. У метанотрофов I типа этот процесс идет по рибулозомонофосфатному пути (РМФ), II типа – по сериновому, а ряд метанотрофов объединен в так называемую группу X, потому что помимо РМФ-пути они используют еще и цикл Кальвина.
Метанотрофы, способные к активной жизнедеятельности при отрицательных температурах, представляют интерес и как модельные объекты для астробиологии, поскольку метан, метанол и формальдегид являются компонентами не только земной криолитосферы. Есть мнение, что такие метанокисляющие микроорганизмы занимают определенное место в гипотетической криптобиосфере Марса (Троценко, Хмеленина, 2008) Различные пути метаболизма отражаются на внутреннем строении клетки. Особенностью метанотрофных бактерий является наличие обширно развитой системы внутрицитоплазматических мембран. У метанотрофов I и Х типа эта система в виде «сложенных стопочек» располагается в центре клетки, а у II типа – по клеточной периферии. Тип расположения служит таксономическим признаком, и в этом смысле ультратонкий срез бактериальной клетки не просто напоминает отпечаток пальца человека, но также, как и он, может использоваться для идентификации. Долгие годы с момента открытия метанокисляющих бактерий в 1905 г. процесс метанотрофии считался строго аэробным, т.?е. идущим только в присутствии свободного кислорода. Однако с 80-х гг. прошлого века ученые стали обнаруживать зоны ассимиляции метана в анаэробных районах. Потребовалось еще много лет, прежде чем было доказано, что в морских осадках этот процесс осуществляется сообществом архей и сульфатредуцирующих бактерий (Boetius et al., 2000). Для пресноводных же осадков определены несколько возможностей. Во-первых, сообществом архей и бактерий-денитрификаторов, восстанавливающих нитраты до нитритов (Raghoebarsing et al., 2006); во-вторых, бактериями-метаногенами, которые при определенных концентрациях метана «переворачивают» свой метаболизм, переключаясь на процесс его окисления (Wang et al., 2014). Однако в лабораторных условиях пока не удалось получить культуры микроорганизмов, способных окислять метан в отсутствии кислорода, и ученые всего мира продолжают заниматься решением этой проблемы. Большинство известных на сегодня метанотрофов обитают в «комфортном» диапазоне температур (20–35?°С). Однако в последнее время появились доказательства существования метанотрофов в активной форме и в экстремальных условиях. Например, разнообразные метанокисляющие организмы были обнаружены в многолетнемерзлых породах Колымской низменности. Предполагается, что даже после длительного (тысячи, миллионы лет) пребывания в вечной мерзлоте метанотрофы остаются способными к активной жизнедеятельности и могут окислять и ассимилировать метан, в том числе при отрицательной температуре (Троценко, Хмеленина, 2008). Среди метанотрофов встречаются и термофильные представители, обитающие при температуре выше 40?°С. Эти микроорганизмы также выделены из очень щелочных (рН > 9,0) и очень кислых (рН < 5,0) сред. Все эти данные позволяют говорить об огромном диапазоне адаптационных возможностей метанотрофов. Метанотрофы на Байкале Озеро Байкал – глубочайшее (максимальная глубина 1637 м) пресноводное озеро в мире – расположено в центре тектонически активной рифтовой зоны. В Среднем и Южном Байкале в конце прошлого века были обнаружены залежи газогидратов, содержащие метан, «законсервированный» в кристаллической решетке из молекул воды (Кузьмин и др., 1998). По сути, все озеро Байкал является резервуаром, в донных осадках которого содержится огромное количество этого газа. Первые исследования метанотрофии на Байкале были проведены в начале 1990-х гг. Б.?Б. Намсараевым (Институт общей экспериментальной биологии СО РАН, Улан-Удэ) и Т.?И. Земской (Лимнологический институт СО РАН, Иркутск). С помощью иммунофлуоресцентного и радиоизотопного метода были получены данные о скорости окисления метана, численности и биоразнообразии метанотрофов в районе выхода гидротемального источника в бухте Фролиха на Северном Байкале (Намсараев, Земская, 2000). В последние годы исследователи заговорили о проблеме увеличения содержания метана в байкальской воде и возможных причинах этого явления (Гранин и др., 2014), и в этом смысле всестороннее изучение биоразнообразия и потенциала микроорганизмов, участвующих в цикле метана, становится актуальной задачей.
Сегодня в лаборатории микробиологии углеводородов ЛИН СО РАН метанотрофия в районах грязевых вулканов, горячих и холодных выходов углеводородов исследуется с помощью комплекса классических микробиологических, а также современных физико-химических и молекулярных методов. В том числе при погружениях на глубоководных обитаемых аппаратах «МИР» в месте выхода газовых гидратов на глубине 1400 м были обнаружены желеобразные микробные маты, состоящие из метанотрофных бактерий, архей и цианобактерий (Zemskaya et al., 2015). Так как вся водная толща озера и верхние 5—10 см донных осадков насыщены кислородом, метанотрофные бактерии обнаруживаются на всех глубинах. Оценить их численность в природном образце позволяет метод флуоресцентной in situ гибридизации (FISH), при котором используются специфические олигонуклеотидные зонды. Наибольшие величины численности и скорости окисления метана были отмечены в придонных зонах грязевых вулканов и в верхних слоях водной толщи (Захаренко и др., 2015; Pimenov et al., 2014). Благодаря использованию метода массового параллельного секвенирования, дающего возможность единовременно анализировать несколько сот тысяч нуклеотидных последовательностей, было обнаружено, что видовой состав метанотрофного сообщества донных осадков и водной толщи различается. Этим методом удалось выявить не только доминирующие микроорганизмы, но и уникальные линии метанотрофов, не имеющие культивируемых гомологов, которые могут быть байкальскими эндемиками.
В метанотрофных микроорганизмах сегодня исследователей привлекает не только их значимость в качестве метанассимилирующей системы, но и богатый «внутренний мир». Речь идет о способности синтезировать полезные продукты, довольствуясь минимальными питательными ресурсами, а также разлагать широкий спектр высокотоксичных соединений. Эти их свойства обусловливают перспективность аэробных метанотрофов для биотехнологий, в том числе технологий биологической очистки. Бактериальную биомассу, полученную «на метане», по аминокислотному составу и содержанию витаминов можно приравнять к рыбной и соевой муке и сухому молоку. А биопротектор эктоин, который накапливается в клетках этих бактерий, даже рассматривают как потенциальное средство терапии диабета и болезни Альцгеймера (Троценко, Хмеленина, 2008). В этом смысле эра изучения байкальских метанотрофов только начинается. Несмотря на то что наши знания постоянно пополняются, а спектр методов, с помощью которых мы сегодня можем исследовать эту очень важную для нашей планеты группу микроорганизмов, расширяется, мы сегодня прикоснулись лишь к самой вершине айсберга, основная часть которого в прямом смысле слова пока еще скрыта в глубоких водах Байкала. Литература Гранин Н.?Г., Верещагина О.?Ф., Козлов В.?В. и др. Изменение концентрации метана в озере Байкал: возможная причина // Рос. конф. «Газовые гидраты в экосистеме Земли 2014», 7—10 апреля 2014 г. Новосибирск: ИНХ СО РАН, 2014. C. 25. Захаренко А.?С., Пименов Н.?В., Иванов В.?Г. и др. Окисление метана в водные толще районы газо- и нефтепроявлений Среднего и Южного Байкала // Микробиология. 2015. Т. 84. С. 98—106. Кузьмин М. И., Калмычков Г. В., Гелетий В. А. и др. Первая находка ГГ в осадочные толще озера Байкал // Докл. РАН. 1998. Т. 362, № 4. С. 541—543. Намсараев Б.?Б., Земская Т.?И. Микробиологические процессы круговорота углерода в донных осадках озера Байкал // Новосибирск: Изд-во СО РАН, филиал «Гео», 2000. 160 с. Троценко Ю.?А., Хмеленина В.?Н. Экстремофильные метанотрофы. Пущино: ОНТИ ПНЦ РАН, 2008. 206 с. Boetius A., Ravenschlag K., Schubert C.?J. et al. // Nature. 2000. V. 407. P. 623—626. Raghoebarsing A.?A., Arjan P. et al. A microbial consortium couples anaerobic methane oxidation to denitrification // Nature. 2006. V. 440. P. 918—921. Zemskaya T.?I., Lomakina A.?V., Shubenkova O.?V. et al. Jelly-like microbial mats over subsurface fields of gas hydrates at the st. Peterburg methane seep (Central Baikal) // Geomicrobiology J. 2015. V. 32. P. 89—100. Wang F.?P., Zhang Y., Chen Y. et al. Methanotrophic archaea possessing diverging methane-oxidizing and electron-transporting pathways // ISME J. 2014. V. 8. P. 1069—1078 Работа выполнена в рамках проекта ФАНО № 0345-2014-0009 Источник: scfh.ru |
 образуют большие скопления над полями залегания байкальских газовых гидратов (фото слева внизу). Общая численность придонного сообщества амфипод в зоне метановой разгрузки «Санкт-Петербург» в 4—6 раз выше, чем в других районах озера. Пищевая цепь этого сообщества включает, помимо криптофитовых водорослей, жгутиковых и цианобактерий, метанотрофные бактерии, благодаря которым метан вовлекается в круговорот углерода.Эпифлюоресцентная микроскопия. По: (Земская, 2010)")

 и II (б) типа отчетливо видны «стопочки» внутрицитоплазматических мембран, расположение которых служит таксономическим признаком. Трансмиссионная электронная микроскопия. Слева – байкальские бактерии-метанотрофы из образца придонной воды. Эпифлуоресцентная микроскопия")
